植物の光感受性変換機構を発見‐LOVドメイン光センサーの光感受性を変換する?(酒井達也教授)
植物の光感受性変換機構を発見‐LOVドメイン光センサーの光感受性を変換する−
【本研究の概要】
本学理学部生物学科・酒井達也教授ら研究グループは、日本工業大学・芳賀健講師とともに植物の光環境認識に働く光センサー「フォトトロピン」(※1)に光感受性の変換をもたらす分子機構が存在することを明らかにしました。本研究成果は米国植物生理学会誌『The Plant Cell』のオンライン版(日本時間4月15日)に掲載されました。
植物の光屈性(※2)は生物が示す最も高感度な光環境応答の一つで、極微弱光の光源を認識し、光源へと成長方向を変化させることができます。一方、植物は太陽のような強い光を発する光源も認識して光屈性を示すこともできます。生物は通常、暗い中では光感受性の高い光センサーを用い、明るい環境では光感受性の低い光センサーを使い分けることによって光強度の大きく異なる環境を認識しています。しかし、同じ光センサーが光感受性を変換して異なる光強度の光を認識するというメカニズムはあまり知られていません。酒井達也教授らは植物の光屈性反応に働く光センサー・フォトトロピン1が極微弱の光から強光まで8桁以上の光強度の異なる光環境で働くことができることに着目し、フォトトロピン1の光感受性のダイナミックレンジ拡張に働く分子機構について研究を行いました。 研究グループは双子葉植物シロイヌナズナ(※3)の光屈性が異常になった突然変異体 rpt2 の光屈性反応を観察しました。その結果、RPT2 タンパク質が発現していない暗所ではフォトトロピン1光センサーは高感受性を示し、光照射によって RPT2 タンパク質が発現するとフォトトロピン1は低感受性に変化することが分かりました。RPT2 は光によって発現が誘導され、フォトトロピン1光センサーに結合することによってその光感受性を下げます。すなわちフォトトロピン1にとって RPT2 はまぶしい光のもとで取り出してつけるサングラスのような働きをする、ということがわかりました。また本研究結果により、「光量と光照射時間によって一次正光屈性と二次正光屈性という異なる光屈性反応が現れるのはなぜか」という植物生理学の古典的問題が解決しました。
この成果により、植物の光センサー・フォトトロピンの光感受性調節機構及びその下流の光屈性誘導機構の解明が進むと考えられます。また、フォトトロピン光感知のコアになる LOV ドメイン(※4)が青色光照射によって構造を変化させることから、光で分子機能をオンオフ可能な分子デバイスとして最近動物細胞の研究によく利用されるようになっており、本研究成果がフォトトロピン分子デバイスの新しい利用法の開発につながると考えられます。
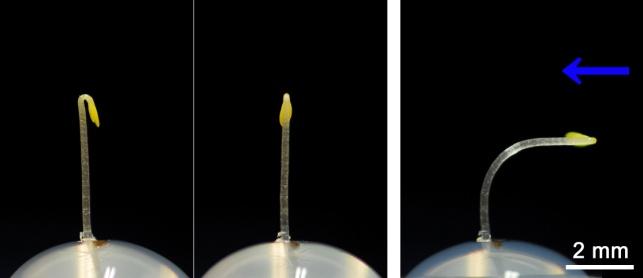
シロイヌナズナ黄化芽生え胚軸の光屈性
横から青色光を照射すると、およそ二時間後には芽生えの胚軸は光源方向へと屈曲する。暗所で育てた黄化芽生えなので、子葉は開いておらずフックが形成されている(左は光照射前に横から見た芽生え、真ん中は光照射前に子葉がある正面側からみた芽生え、右は矢印方向から3時間青色光を照射後の芽生えの様子)。
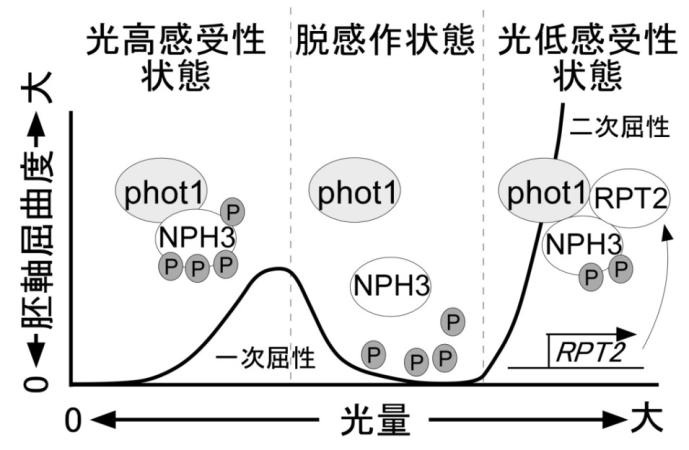
本研究で明らかになった光屈性誘導機構のモデル図照
照射する青色光の光量が低い芽生えにおいては(左)、フォトトロピン1(phot1)は光高感受性を示す。このときリン酸化されたシグナル伝達因子 NPH3 とフォトトロピンは複合体を形成し、光屈性誘導のシグナルを流して芽生え胚軸の屈曲が誘導される。このような光環境ではRPT2はほとんど発現していない。光量が増えると phot1 活性化に伴った NPH3の脱リン酸化がおきて、phot1とNPH3の結合が解消し、光屈性誘導のシグナルが流れず胚軸屈曲も現れない脱感作状態が生じる(中央)。照射時間が長くなり光量が増加すると RPT2 の発現が誘導され、phot1 は RPT2 と結合して光低感受性状態に変換する(右)。phot1が光低感受性状態に変換することによって、NPH3 の脱リン酸化反応は緩和され、phot1とNPH3は再び結合して光屈性誘導のシグナル伝達がおきる。脱感作状態をはさんで低光量でおきる光屈性を一次正光屈性、高光量でおきる反応を二次正光屈性とよぶ。
イメージイラスト
暗所においては植物の黄化芽生えは光源方向を識別する光感受性が極めて高い(左)。明所においては、RPT2 タンパク質を発現し、光感受性を下げて光源方向を識別できるようにし(光屈性寛容)、成長方向を調節する(右)。

【用語解説】
- (※1) フォトトロピン
緑藻類から高等植物まで広く保存された植物に特異的に存在する青色光受容体で、光屈性、葉緑体光定位運動、気孔開口運動などの植物の光環境応答に働く。2つのLOVドメインに1つのタンパク質リン酸化酵素ドメインをもち、2つのLOVドメインにはそれぞれ青色光を吸収するフラビンモノヌクレオチドが結合している。LOVドメインにおける青色光吸収がフォトトロピン光受容体の構造を変化させて、細胞内で一連のシグナル伝達がおきると考えられている。 - (※2) 光屈性
胚軸や茎の成長方向を光源方向に向けることによって、より光を吸収し光合成活性を高めるために働く植物の光環境応答。19世紀にダーウィンによって観察・記述され、20世紀における植物ホルモン・オーキシンの発見につながった。光照射方向の情報がどのようにオーキシンの不均等勾配形成を促し、偏差成長による軸器官の屈曲に至るのかについては未だ明らかになっていない。 - (※3) シロイヌナズナ
学名はArabidopsis thaliana。アブラナ科シロイヌナズナ属の1年草。2000年にモデル実験植物として植物で初めてゲノム塩基配列が完全に解読された。 - (※4) LOVドメイン
LOV は light、oxygen、voltage の略。細菌、古細菌、原生生物、植物、真菌で、光、酸素、電圧の感知に働くタンパク質によく保存されたアミノ酸配列であったことから名付けられたタンパク質ドメイン。フォトトロピンの LOV ドメインはフラビンモノヌクレオチドと非共有結合して存在しており青色光を吸収するとその構造が変化することから、最近、動物細胞の研究などによく用いられている。
(例;http://www.nature.com/nature/journal/v518/n7537/full/nature14128.html)
お問合わせ先
- 新潟大学理学部生物学科
教授 酒井達也
TEL/FAX:025-262-7880
※酒井達也教授のプロフィールはこちらです。